 |
||||
| |
||||
| |
||||
| |
||||
![]()
![]()
![]()
![]()
![]()
![]()
| 微小管制御を介した細胞極性・生体秩序形成維持機構の解明 と、その異常に起因するヒト脳疾患の発症メカニズムの解明 |
|
|
|
|
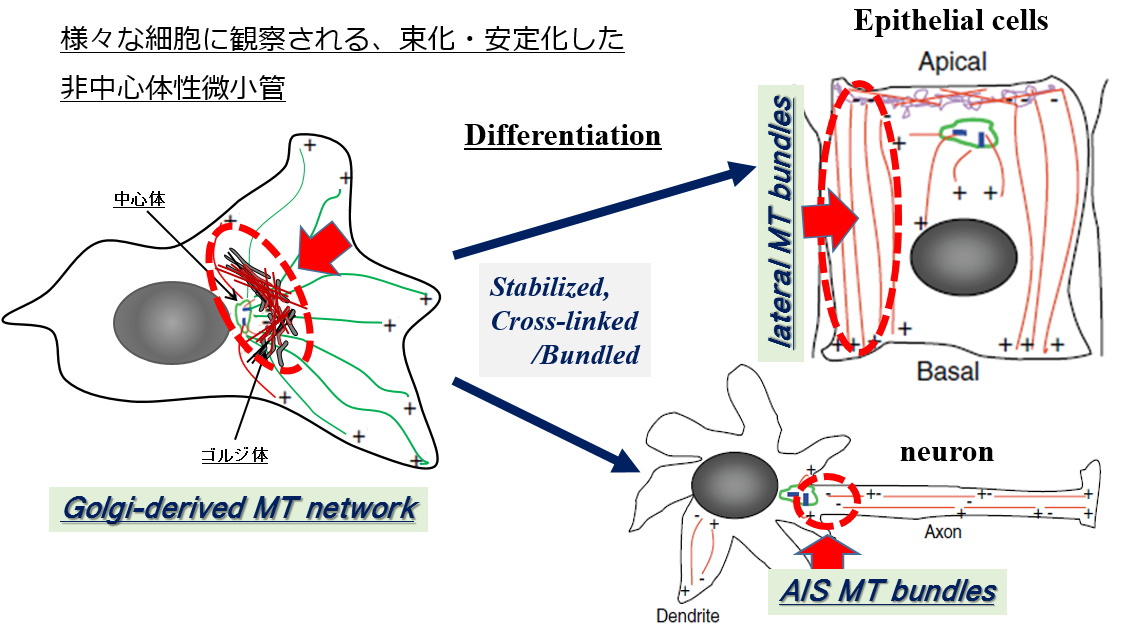
| [1] 微小管束化・安定化因子MTCLタンパク質による生体機能制御の研究 非対称性・極性を発達させた分化した細胞のほとんどは、中心体から解離した微小管が束化・安定化して、細胞固有の配向(細胞内での走行、並び方)を示しています。そして、これらの細胞特異的な配向を示す微小管が、それぞれの細胞の形態や機能の発達に不可欠な役割を果たしています。しかし、こうした微小管の配向を制御する仕組みは近年まであまり分かっていませんでした。 |
|
 |
|
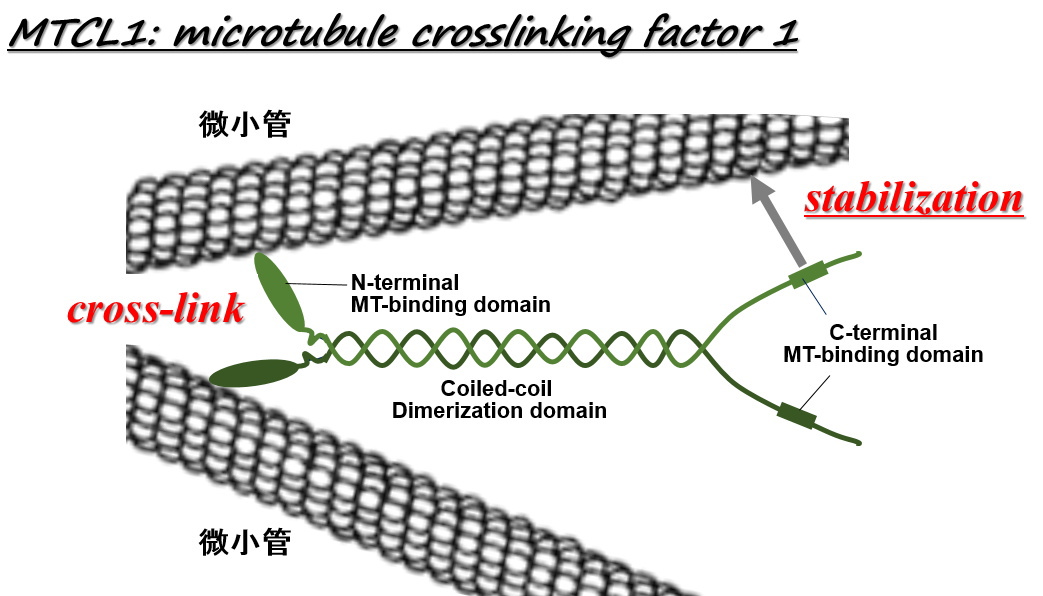
| こうした中で我々は、非中心体性微小管の束化・安定化を制御する微小管制御因子を世界で初めて発見し、MTCL1 (microtubule crosslinking factor 1)と命名しました(J. Cell Sci. 2013, Nat. Communi. 2014)。 | |
 |
|
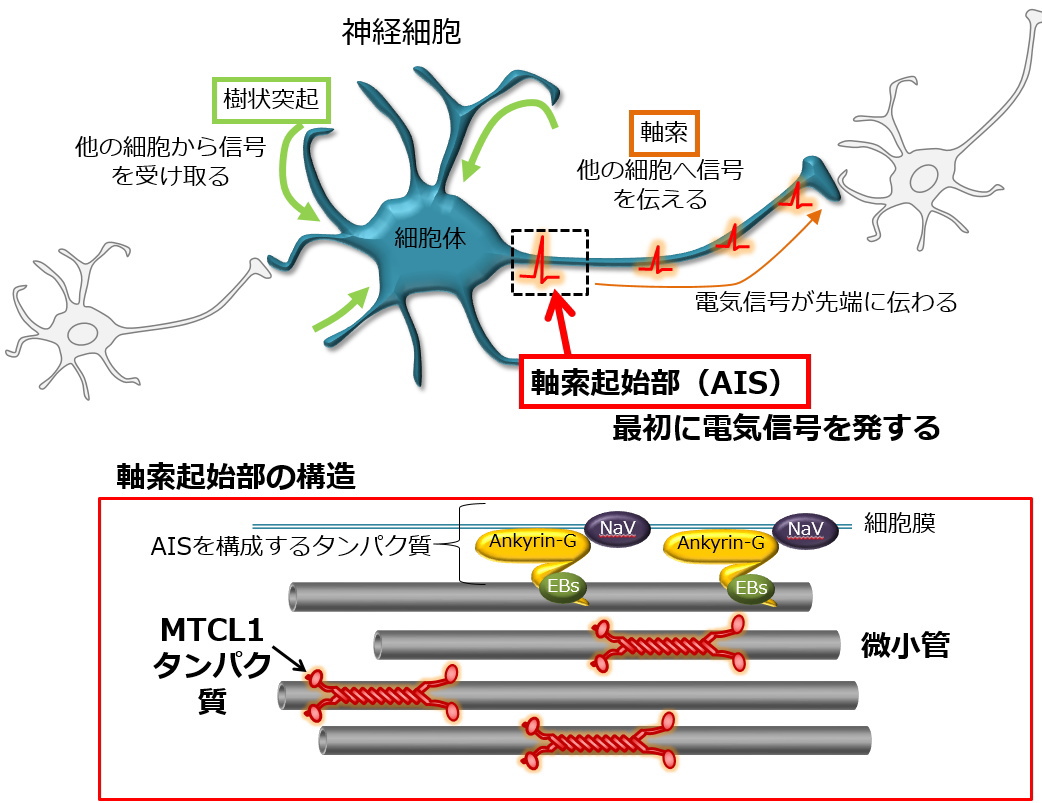
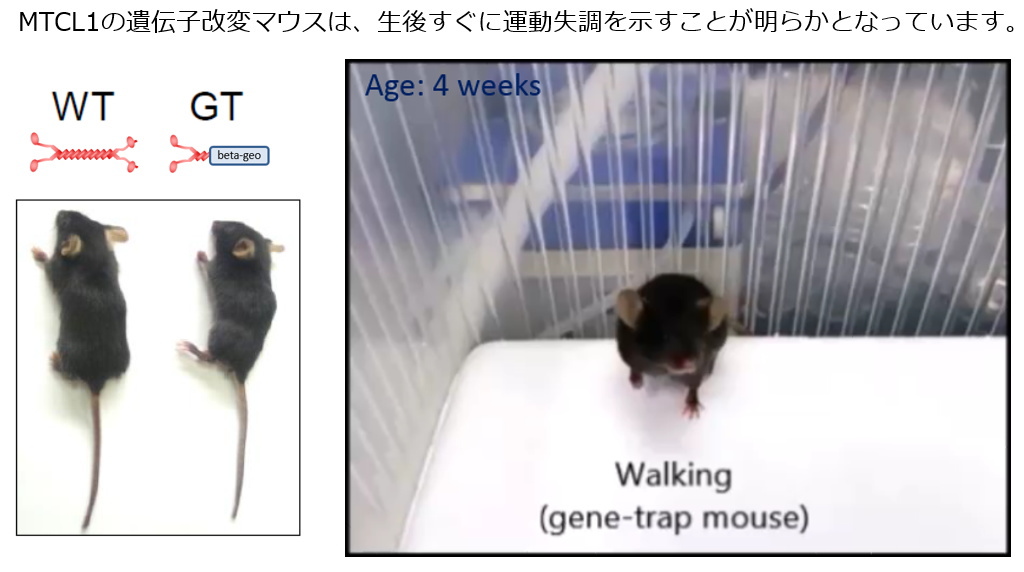
| マウス個体を用いた研究からは、このMTCL1による微小管束化・安定化活性は、特に小脳プルキンエ細胞の「軸索起始部(AIS)」の形成・維持に不可欠であることが示されました。神経細胞の軸索の根元に発達するこのAISの存在は、神経細胞の極性の発達(入力と出力を分担する樹状突起と軸索の発達)と、軸索を通じて他の細胞に信号を伝える活動電位の発生に不可欠な役割をしています。従って、正常なMTCL1を失ったマウス(遺伝子改変マウス)の小脳プルキンエ細胞は正常に働くことができず、そのためマウスはヒト脊髄小脳変性症に類似した小脳の変性と運動失調を示すことが判明しました。(EMBO J. 2017) | |
 |
|
 |
|
| 現在は、こうしたMTCL1の研究をさらに発展させるため、様々な手法を駆使して以下のような研究を精力的に展開しています。 ① MTCL1分子の構造機能相関の研究 -- ex. MTCL1のゴルジ体局在領域の同定 ② MTCL1のC末端領域が微小管の重合状態を安定化する機構の解明 ③ MTCL1が神経細胞軸索起始部構造の発達を促進する分子機構の解明 ④ MTCL1の他の臓器(特に発現の高い、肺・卵巣等)における機能の解析 * MTCL1遺伝子の多型は肺気腫の危険因子であることが最近、報告されています。 ⑤ MTCL1のパラログ分子、MTCL2の機能解析(遺伝子改変マウスの解析を含む) ⑥ MTCL1, 2の細胞分裂における機能 --> リン酸化を介した機能制御機構の解明 |
|
|
|
|
| [2] チューブリン特異的シャペロンTBCDの解析を通じた生後脳の発達機構の解明 | |
|
|
|
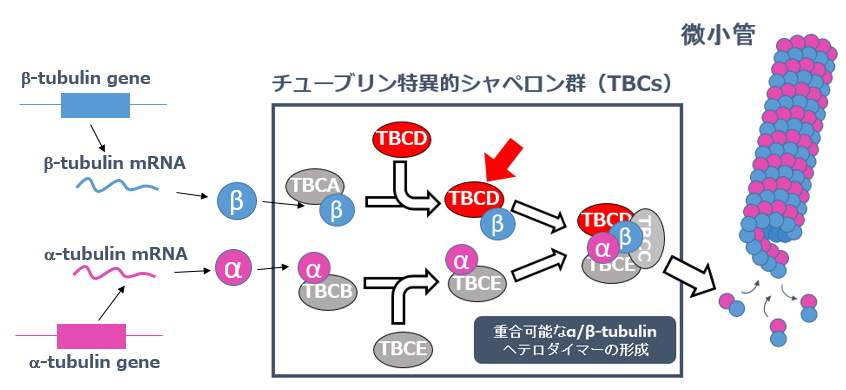
| 微小管を構成するα、β-tubulinが正常に折りたたまれて重合可能なヘテロ2量体を形成するには、専用のシャペロンシステムが必要です。 | |
 |
|
驚いたことに、このシャペロン系を構成するTBCDの遺伝子に点変異が生じると、ヒト幼児の生後の脳の発達異常が引き起こされることが2016年に報告されました(ex. Miyake et al. Am. J. Hum. Genet. 2016)。 私達の研究室では、もう一つの研究課題としてこの「早発性神経変性脳症」と名づけられた脳遺伝性疾患の発症機構の解明を目指しています。そしてこの研究を通じて、いまだ解析の遅れている「生後の脳構造の発達機構の解明」に切り込もうとしています。 |
|
 |
|
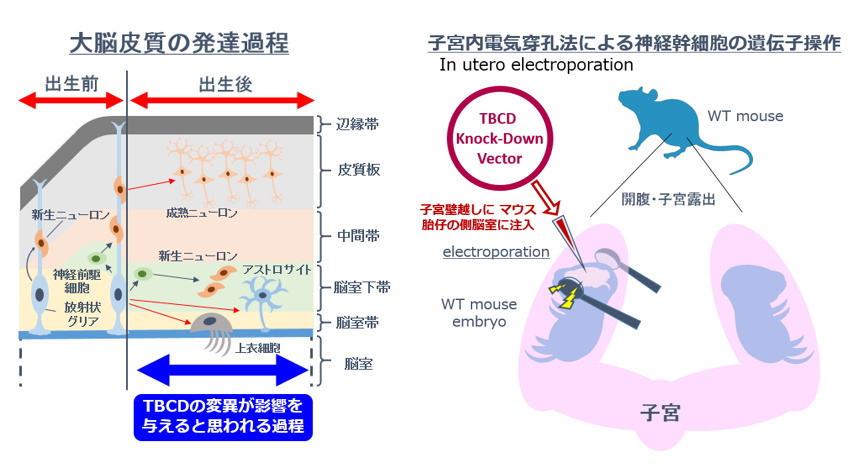
| 近年、大脳皮質層構造の形成機構の解明が大きく進みましたが、それは「子宮内穿孔法(in utero electroporation法」という遺伝子導入技術が発達し、胎仔の
脳室に面して存在する神経幹細胞に特異的に系譜マーカーやノックダウンベクターなどを導入できるようになったことに大きく依存しています(上右図)。ただ、この方法で導入されたベクターの幹細胞内の濃度は分裂に応じて薄まってしまうため、胎仔期に導入したベクターを長期にわたって維持し、生後に見られる現象の解析に利用することができないという技術的困難が残っていました。このため、神経細胞の産生・移動による基本的大脳皮質層構造の形成が進んだ後、出生直前から急速に進む種々のグリア細胞の産生や、神経細胞の樹状突起の複雑化、シナプスの形成など、脳体積の急速な増大、脳構造の本格的な発達を引き起こすさまざまな重要なプロセスの解明がいまだに大きく遅れています。 TBCDの変異が影響する過程はこの生後脳の発達過程と考えられるため、我々の研究室ではこの難点を解決を試みてきました。そして最近、当研究室の特任助教が生後脳の解析を可能とする技術的開発に成功しました。 現在、この技術の洗練化を進めることを通じてTBCD変異が影響する過程を明らかにすることを目指しています。またその分子機構に迫るため、TBCDの機能の細胞生物学的解析を並行して進めています。 |
|