|
|
|
発生・分化・形態形成などの複雑な生命現象は特定の遺伝的プログラムに従って正確に行われる。これらのプログラムは主に転写段階で調節されながら進行することがすでに知られており、転写制御機構の理解はこれらの生命現象を解き明かす上で非常に重要であることはいうまでもない。しかしながら千種類以上の転写調節因子が単離された現在でも転写調節の分子機構に関する知見は極めて限られている。遺伝子特異的に作用するこれらの転写調節因子は、蛋白質間相互作用を介して転写開始点上に形成される基本転写装置の数あるいはその活性を制御し、各遺伝子の発現量を規定している。一方標的となる基本転写装置は多数の基本転写因子(TFIIA,
B, D, E, F,H)とRNAポリメラーゼ II から構成され、遺伝子の種類を問わず機能する。残念ながら現時点では転写調節因子と基本転写装置の機能的相互作用の実体は不明であり、シグナルの送り手である転写活性化ドメインに要求される生化学的な性質、あるいはシグナルの受け手である基本転写装置内部に存在する活性化に必要な分子スイッチの構造といった基本的な問題にすら明確には答えられないのが実状である。また最近になって転写調節に関与する普遍的転写因子群が次々に単離・同定され、転写調節の分子機構を統一的に理解することはますます困難になりつつあるように見える。そこで我々の研究室では、転写調節において極めて重要な役割を果たす基本転写因子TFIIDに注目して研究を進め、一見混沌としてみえる現在の状況を整理し、転写調節の分子基盤を明確に説明し得るコンセプトの確立を目指している。
2. 基本転写因子TFIIDに内蔵された分子スイッチTANDの機能 基本転写因子のなかでもTATAボックス/イニシエーター/DPE等のプロモーター構造を認識するTFIIDは、転写開始前複合体のアッセンブリーに際して核となる分子であり、転写調節因子から受け取った信号を転写量の増減へと変換するうえで中心的な役割を果たす。我々はショウジョウバエ胚の抽出液からTFIIDを単離し、約10個近い構成成分を同定するとともに、TATAボックス結合タンパク質(TBP)以外の他のTFIIDサブユニット(TAFs)が転写調節に必須の役割を果たすことを示した(JBC,vol.268,p17554,1993)。また各サブユニットをコードするcDNAをショウジョウバエより単離し(PNAS,vol.90,p5896,1993/MCB,vol.13,p7859,1993/Nature,vol.367,p484,1994)、これらの機能を解析する過程において、最大サブユニットdTAF230(現在はdTAF1と呼ばれる)のN末端にTBPと強く結合し、その機能を阻害する新規の活性が存在することを見出した(Genes & Dev,vol.7,p1033,1993/PNAS,vol.91,p3520,1994)。この阻害活性は酵母からヒトまで種を越えて強く保存されていたことから、遺伝学的解析の容易な酵母を新たに材料に加え、この阻害活性領域(TAF N-terminal domain:以下TANDと略)について詳細な解析を行い(MCB,vol.18,p1003,1998/JBC,vol.273,p32254,1998)、TANDとTBPの相互作用の変化こそが転写調節因子によるTFIID活性化の鍵を握る初発段階の反応ではないかと考えるに至った(PNAS,vol.97,p7178,2000)。現在はこれまでの知見を説明し得る転写活性化の分子モデル(二段階ハンドオフモデル)を新たに構築し、その妥当性について様々な角度から検証を進めている。
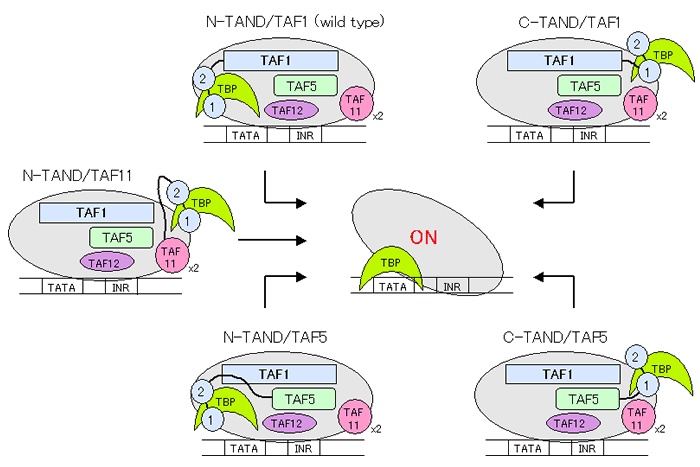
我々は、酸性型転写活性化ドメイン(AD)とTANDのN末端側サブドメイン(TAND1)の間に成立する機能的互換性を見出し、ADはTBP上でまずTAND1と置き換わることにより転写活性化の引き金を引くのではないかという上記二段階ハンドオフモデルを提唱するに至った。しかしこのモデルを別の観点から見ると、TANDの役割はTATAボックス近傍に不活性型状態のTBPを濃縮すること、及びADからのシグナルに適切に応答してTBPをTATAボックスに送り込むことであると理解することもできる。すなわちTAND-TBP複合体はADと必要な相互作用ができれば良いのであって、他のTAFsとは独立に機能する(自律的な機能を有する)分子デバイスと捉えることも可能である。実際にTANDをTAF1のN末端からC末端へ、TAF5のN, C両末端へ、TAF11のN末端へと移植したところ、TANDはTFIID内部の位置に関係なく正常に機能し得ることが明らかとなった(MCB,vol.24,p3089,2004)。従ってTANDは必ずしもTAF1のN末端に存在する必要はなく、ADからのシグナルを受け取ることができさえすれば、機能を保持したままTFIID内部を自由に動き回れるものと考えられる。
2-2. 新規TBP結合ドメインであるTAND3の同定とその機能解析 これまで出芽酵母のTANDはTAND1(10-37aa)とTAND2(46-71aa)という二個のサブドメインのみから構成されると考えられていたが、最近我々はTAND2のC末端側直近に転写活性化能を有する領域が存在することを新たに見出し、その最小活性領域を決定してTAND3(82-139aa)と名付けた。TAND1やTAND2と同様、TAND3もTBPに結合する活性を有しており、TBPの変異体解析からTAND1とTAND3はともにTBPのDNA結合表面側を一部重複するような形で認識することが明らかとなった。興味深いことに、複数の遺伝子がTAND3の機能に依存して転写されることから、これらの遺伝子のプロモーター上ではTAND3によるTBP機能の抑制がADによって解除されることで転写が開始されるものと考えられる(JBC,vol.278,p45888,2003)。
2-3. 転写/翻訳開始点シフトによるTAND欠失型TAF1タンパク質の発現制御 出芽酵母とショウジョウバエ間のキメラ型TANDを有するTAF1遺伝子を作成し、その性質を調べた。ショウジョウバエ由来のTANDは出芽酵母の細胞内で正常に機能できず、上記キメラ型遺伝子を発現させた場合にはTAND欠失型TAF1タンパク質が著量蓄積することが明らかとなった。驚くべきことに、この蓄積はTAF1遺伝子の転写開始点が下流にシフトしたことに伴い、翻訳開始点がTAND以降のメチオニン残基にシフトしたためであることが示された(Genes to Cells,vol.9,p709,2004)。野生株においても同様の転写/翻訳開始点シフトによってTAND(1+2) or TAND(1+2+3)欠失型TAF1タンパク質がわずかながら生産されることから、異なるTAND領域を欠くTFIIDアイソフォームによって制御される特異的な標的遺伝子が存在する可能性なども考えられる。
2-4. TAND欠失型TAF1遺伝子と合成致死性を示すNSL遺伝子群の単離と解析 TAND領域が転写制御において果たす役割を遺伝学的な見地から明らかにするため、TAND欠失型TAF1遺伝子(taf1-deltaTAND)に対して合成致死性を示すNSL(TAND synthetic lethal)遺伝子変異株のスクリーニングを行った。これまでに15株を単離し、2株はTBP遺伝子変異株(spt15-S118L,P65S)、1株はTAF12遺伝子変異株(taf12-L420S)であることを明らかにした(JBC,vol.276,p395,2001/NAR,vol.31,p1261,2003)。またspt15-118L,p65S変異株はポストTBPリクルートメントステップ、taf12-L420S変異株はプレTBPリクルートメントステップの欠損を示し、いくつかの標的遺伝子の転写を調べたところ、taf1-deltaTAND変異株は後者とよく似た表現型を示すことが明らかとなった。この結果は、TANDがTBPリクルートメントステップに関与するという上記二段階ハンドオフモデルの妥当性を間接的に支持するものと考えられる。残りの12株についても原因遺伝子の特定を終了し、現在その性質について詳しい解析を進めている。
出芽酵母のTFIIDサブユニットの一つであるTAF1遺伝子について全長にわたって点変異をランダムに導入することにより多数の温度感受性変異株を単離し、TAF1と遺伝学的に相互作用する分子について同定を進めている。また得られた二種類の点突然変異株(N568delta,T657K)については独自に開発したin vivo転写系を用いて解析を進め、RPS5,TUB2遺伝子等のTATA-less コアプロモーターの認識異常、コアプロモーターと転写調節因子の組み合わせに依存した転写活性化能の欠損、コアプロモーターの種類によるTATAボックス挿入効果の違いなどをすでに明らかにしている(MCB,vol.20,p2385,2000/JBC,vol.276,p25715,2001)。現在はこれまでに同定した複数の標的遺伝子のコアプロモーターについて詳細なキメラ解析を行い、TAF1がその認識に関与すると考えられるコアプロモーター中の分子的特徴をヌクレオチドレベルで明らかにするべくさらに解析を進めている。
酵母細胞内においてTAND領域(1-208 aa)と相互作用する因子の探索(TANDをbaitとするcytotrap screening)を行い、酵母のHMGBタンパク質の一種であるHMO1を同定した(MCB, vol.27, p6686, 2007/NAR, vol.36, p1343, 2008)。生化学的な解析の結果、HMO1は直接TBPと相互作用し、TFIIDと共精製されることが明らかとなった。HMO1遺伝子破壊(hmo1Δ)株では複数の遺伝子の転写量の変化と生育遅延が見られ、hmo1ΔとTAND欠失変異(taf1ΔTAND)を組み合わせた二重変異株では、hmo1Δ単独変異株に比べて、より顕著な生育阻害が見られた。またHMO1タンパク質の過剰発現はTAND1欠失株の生育を特異的に阻害した。以上の結果は、HMO1がTAND/TBP/TFIIDとともに働く転写因子であることを示している。
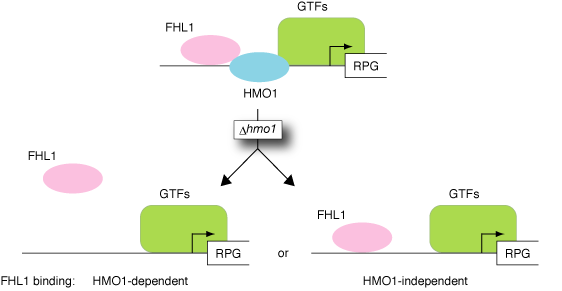
4-1. ゲノム上のHMO1結合部位の同定と転写及び各種転写因子のリクルートメントにおけるHMO1依存性 HMO1は、リボソームRNA(rRNA: ribosomal RNA)遺伝子(プロモーター+コーディング領域)及びリボソームタンパク質(RP: Ribosomal Protein)遺伝子(プロモーター領域)に結合し、RP遺伝子群の主要な転写調節因子であるFHL1のRPプロモーター結合において必須の役割を果たす。我々は、詳細なChIP解析を行い、HMO1が35S rRNA遺伝子のコーディング領域全体にRNAポリメラーゼI依存的に結合すること、またHMO1は多くのRPプロモーターに結合するが、その結合量は遺伝子ごとに大きく異なることを見出した(MCB, vol.27, p6686, 2007)。興味深いことに、FHL1は、HMO1結合量の多いものにはHMO1依存的に、またHMO1結合量の少ない(あるいは結合しない)ものにはHMO1非依存的に結合しており、これらの結合特性はプロモーター配列によって規定されることが明らかとなった。またゲノムワイドなChIP-chip解析により(東京大学・白髭克彦教授との共同研究)、RP遺伝子群(計138個)は、HMO1の結合量、FHL1/RAP1の結合の有無及びそのHMO1依存性から計13種類のサブグループに分類できることが示された。HMO1遺伝子破壊の転写量への影響もRP遺伝子ごとに大きく異なっていたことから、従来RP遺伝子群は一様な制御を受けるとされてきたが、実際にはHMO1/FHL1/RAP1など複数の転写調節因子によって多様な制御を受けているものと考えられる。
hmo1Δ変異は、TBP変異やTFIIA変異と組み合わせることにより、酵母の生育において負の合成効果を示すが、逆に一部のTFIIB変異による生育阻害に対しては回復効果を示す(NAR, vol.36, p1343, 2008)。これらのTFIIB変異株では転写開始部位(TSS: transcriptional start site)が下流側にシフトすることから、hmo1Δ変異のTSSに及ぼす影響を調べた。その結果、hmo1Δ変異株では、HMO1標的遺伝子特異的にTSSが上流側にシフトすることが明らかとなった(NAR, vol.36, p1343, 2008)。現在、HMO1がTSSを決定する分子機構について、さらに解析を進めている。
5. メディエーター(ミドルモジュール)の構成成分であるMed9の変異体解析 出芽酵母のメディエーターは、ヘッド(Med6, Med8, Med11, Med17, Med18, Med20, Med22)、ミドル(Med1, Med4, Med7, Med9, Med10, Med19, Med21, Med31)、テール(Med2, Med3, Med5, Med14, Med15, Med16)、Cyc-C(Med12, Med13, Cdk8, CycC)の計4モジュールから構成される巨大なタンパク質複合体であり、TFIIDとともに真核細胞の転写制御において重要な役割を果たす。我々はHIS4遺伝子の転写制御におけるTANDの役割との類似性から、ミドルモジュールの構成成分であるMed9に注目し、その変異体解析を行った(Genes to Cells, vol.14, p53, 2009)。その結果、Med9は種特異的なN末端側ドメイン(Med9-N: 1-63 aa)と種間保存性の高いC末端側ドメイン(Med9-C: 64-149 aa)から構成されること、及びMed4/Med7との結合やミドルモジュールへの組み込み、in vitroにおける転写活性化能などの主な機能はMed9-Cによって担われていることが明らかとなった。またmed9 (Δ119-149 aa) 株由来の細胞破砕液(WCE)は、野生株由来のWCEから生化学的にメディエーターを除去し、さらにそこにMed9 (Δ119-149 aa) を含む変異型メディエーターを戻したものとは、TBP/Taf11のリクルートメントにおいて異なる挙動を示したことから、med9株由来のWCE中にはTFIIDのプロモーター結合に影響を与える未知の因子が含まれているものと考えられる。現在、この因子の精製・同定を進めている。
6. TFIIDサブユニットのゲノムワイドなプロモーター結合解析 出芽酵母のクラスIIプロモーターにおけるTBP結合は、5個のサブユニットを共有する二種類の複合体であるTFIID(15サブユニット)とSAGA(20サブユニット)によって制御されている。DNAチップを用いた発現解析の結果から、TFIIDは主にハウスキーピング型遺伝子群(TATA-lessプロモーターを持つことが多い)を、またSAGAは主にストレス誘導性遺伝子群(TATA-containing プロモーターを持つことが多い)を担当すると考えられている。我々はTFIIDの全Tafサブユニット(Taf1-14), SAGA (Gcn5), NC2 (Bur6/Ncb2), TFIIB (Sua7), TFIIE (Tfa2), TFIIF (Tfg1), TFIIH (Tfb3), RNAポリメラーゼII (Rpb1)についてゲノムワイドなプロモーター結合解析を行い(東京大学・白髭克彦教授との共同研究)、野生株とtaf1-T657K株において得られた結果を相互に比較することにより、これら基本転写因子群の新たな機能の解明を試みた(NAR, vol.38, p1805, 2010)。 TFIIBとNC2は、結晶構造解析の結果からプロモーターに同時に結合することはないと考えられてきたが、本研究において両者は極めてよく似た結合プロファイルを示すことが明らかとなった。一方、taf1-T657K変異は、RP遺伝子プロモーターの場合、それ自身の結合にはあまり影響を与えないが、TBP結合や転写開始前複合体形成(特にTFIIBの取り込み以降のステップ)を阻害すること(ただしSAGAのリクルートには影響を与えない)、またゲノムワイドに見た場合にはTaf2-14のうちTaf2のDNA結合量のみを特異的に増加させることなども明らかとなった。さらにTaf1-14のプロモーター結合プロファイルをゲノムワイドあるいは個別の遺伝子ごとに詳細に比較することにより、TFIIDのコンフォーメーションはプロモーターごとに異なる可能性が示唆された。また連続的なクロマチン免疫沈降実験(sequential ChIP)により、TFIIDとSAGAがRP遺伝子プロモーターに同時に結合することも初めて明らかとなった。
7. TATA-less RPS5プロモーター中に存在するコアプロモーターエレメントの同定 RPS5遺伝子はTATA-lessプロモーターを有しており、TFIID依存的に転写される(MCB, vol.20, p2385, 2000/JBC, vol.276, p25715, 2001)。出芽酵母ではTATAボックス以外のコアプロモーターエレメントが同定されておらず、TFIIDが認識するTATA-lessプロモーター中のシス配列については明らかにされていない。そこで我々はRPS5プロモーター中のコアプロモーターエレメントを同定するため、プライマー伸長法を用いて詳細なシス配列解析を行った(NAR, in press, 2010)。その結果、TSSの20〜90 bp上流に存在するAAAA, TTTT, AATAなどの短いATストレッチ(計9箇所)がコアプロモーターエレメントとして独立かつ協調的に機能することが明らかとなった。また出芽酵母のコアプロモーターエレメントは、高等真核細胞の場合とは異なり、コア因子(TBP, TFIID, SAGAなど直接コアプロモーターエレメントを認識する転写因子)の結合に関与する配列CE(core factor binding element)とproductiveな転写開始を可能とする配列IE(initiation element)に分類されること、ならびにCE上に形成された転写開始前複合体からRNAポリメラーゼIIが下流側のIEまでスキャンすることにより転写が開始することなどが示された(高等真核細胞ではCEとIEが重複して存在することが多い)。現在、他のTATA-less promoterについても同様の解析を進め、その一般性について検証を行っている。
8. CLN2遺伝子の発現を制御する多機能性因子Ssd1の機能解析 出芽酵母には由来の異なる株が複数存在し、特定遺伝子のアレルの違いに起因する表現型がいくつか報告されている。細胞周期の進行、細胞壁の維持、寿命の決定など様々な生命現象への関与が示唆されるSSD1には、SSD1-V (functional) とssd1-d (non-functional) の二種類のアレルが存在し、両者は異なる表現型を示すが、Ssd1自身の分子機能については不明な点が多い。G1期サイクリンをコードするCLN2は、taf1-N568Δ株において転写が低下する遺伝子の一つである(MCB, vol.20, p2385, 2000)。制限温度下(37.5℃)においてtaf1-N568Δ株を培養すると、ssd1-dアレルを有する場合(taf1-N568Δ ssd1-d株)のみCLN2の発現が消失することから、CLN2はTaf1とSsd1の両因子により制御されていることが明らかとなった。また制限温度下においてtaf1-N568Δ SSD1-V株を培養し、フェナンスロリン投与(転写阻害)後のCLN2 mRNA量の変化を経時的に観察したところ、CLN2は転写活性化とmRNA安定化の両側面からSsd1により制御されていることが明らかとなった(Genes to Cells, in press, 2010)。Ssd1の両機能に必要なCLN2上のシス配列を探索したところ、5’-UTRが双方に必須であるという興味深い結果が得られた。またSsd1はこの領域を含むmRNAに対して強く結合し、その翻訳を阻害することが示唆された。近年6種類のタンパク質群(Cbk1, Kic1, Mob2, Hym1, Tao3, Sog2)から構成されるRAM(Regulation of Ace2 activity and cellular Morphogenesis)シグナルネットワークに含まれるCbk1キナーゼがSsd1のN末端領域をリン酸化することにより、Ssd1による翻訳阻害を解除するというモデルが提唱されている。このモデルではSsd1と細胞壁関連タンパク質をコードするmRNAの関係について議論しているが、CLN2 mRNAに対しても同様の局所的な翻訳制御システムが働くものと考えられる。現在、この点に関してさらに検証を進めている。
9. バフンウニを用いた初期胚発生における転写活性化機構の解析 バフンウニの胚発生において重要な役割を果たすOtx遺伝子は互いに異なるプロモーターにより支配される二種類のタンパク質(初期型HpOtxEと後期型HpOtxL)をコードする。HpOtxEはTATA-lessプロモーター、HpOtxLはコンセンサスなTATAボックスを含むプロモーターにより制御されており、それぞれの発現は未孵化胞胚期、孵化胞胚期に始まる。これらのプロモーターの発現時期特異性におけるTATAボックスの寄与を調べるために、種々のプロモーターコンストラクトを作成してウニ胚への遺伝子導入実験を行った。レポーター遺伝子として用いたルシフェラーゼの活性を指標に発現時期を調べたところ、HpOtxLプロモーターの場合にはTATAボックスの欠失により発現時期が早まり、逆にHpOtxEプロモーターの場合にはTATAボックスの挿入により発現時期が遅れることが明らかとなった。より感度の高いRT-PCR法を用いて直接レポーター遺伝子のmRNA量を定量した場合にも同様の結果が得られたことから、Otx遺伝子の発現時期の決定にはTATAボックスの有無が重要な役割を果たすものと考えられる(NAR,vol.30,p3034,2002)。
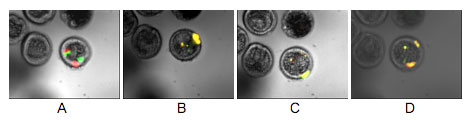
図1. 同一胚を経時的に顕微鏡観察して得られた蛍光画像.
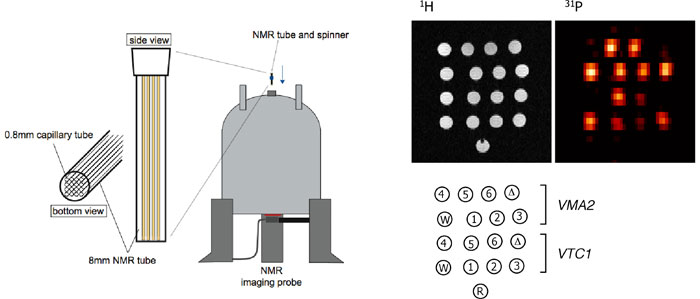
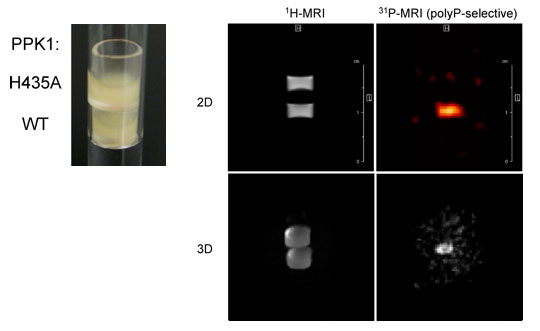
本研究では、生物個体における遺伝子発現を非破壊的に計測する新規手法の開発を行い、その手法を用いて真核細胞の転写制御を支える分子的基盤及びその作動原理を明らかにする。特に新規レポーター遺伝子の開発とMRI計測手法の改良を重点的に進め、これまで不可能とされてきた不透明な生物個体の深部組織における遺伝子発現を細胞レベルの解像度でリアルタイムに可視化することを目指している(京都大学工学研究科・白川昌宏教授との共同研究)。 10-1. 遺伝子発現の可視化を可能とする新規レポーター遺伝子の探索 本研究では、外来基質を一切投与することなく遺伝子発現を高感度かつリアルタイムにモニタリングし得る非侵襲的な新規計測技術の開発を目標としている。我々は、全ての生物に存在するポリリン酸を31P-NMRを用いて可視化することが最も有効な方法論であろうと考え、ポリリン酸の蓄積を定量的に誘導できるレポーター遺伝子の探索と、高解像度でポリリン酸の蓄積を検出できるイメージング技術の確立を目指して、以下の研究を行った。 10-2. ポリリン酸の蓄積を定量的に可視化するイメージング技術の開発(出芽酵母) 出芽酵母細胞内の遺伝子発現レベルを非侵襲的・定量的にモニタリングし得る上記レポーター遺伝子を利用し、多種類の酵母株を同時に測定することができるイメージング手法の開発を行った。まずφ8 mmの直径を持つNMR測定管内に酵母のコロニーを効率よく配置するための器具を複数考案し、これらの器具を用いて測定管内に野生株、Δvtc1株のコロニーを配置した。化学シフト選択的な31P-NMRシグナルによるイメージング手法(31P-MRI, CHESS)を適用することにより、ポリリン酸のシグナルのみを極めて高いコントラストで可視化することに成功した。さらに発現強度の異なる6種類のプロモーターの支配下にVMA2, VTC1レポーター遺伝子を組み込んだ酵母株、及びそのコントロール株(野生株, Δvma2株, Δvtc1株)をそれぞれφ0.8 mmの極細キャピラリー管に移送し、計16種類の酵母株全てを同時に測定する方法論の開発にも成功した(図1)。この方法で測定したポリリン酸蓄積量とmRNA量との間にはやはり強い相関が見られたことから、出芽酵母細胞における遺伝子発現を非侵襲的・定量的にイメージングするための基盤技術として、本システムは極めて有用と考えられる(NAR, vol.34, e51, 2006)。 10-3. ポリリン酸の蓄積を定量的に可視化するイメージング技術の開発(動物細胞) VTC遺伝子群は出芽酵母に特異的であり、高等真核生物にはオルソログが存在しない。一方、VMA遺伝子群の場合には明らかなオルソログが認められるものの、これらの遺伝子を欠くマウスは胚性致死となることが知られている。そこで原核細胞に広く分布するポリリン酸合成酵素であるポリリン酸キナーゼ(PPK1)に着目し、動物培養細胞における本酵素の発現とポリリン酸の蓄積量について検討を行った。その結果、複数の動物培養細胞において大腸菌由来の活性型PPK1遺伝子を発現させた場合にのみポリリン酸の蓄積が認められ、その細胞当たりの蓄積量はほぼ出芽酵母細胞と同等(~120mM)であることが示された。また薬剤誘導性プロモーターの下流にPPK1遺伝子を連結し、ルシフェラーゼとの比較を行ったところ、両者はほぼ同等の検出感度を有することが明らかとなった。蓄積したポリリン酸を指標とするPPK1発現細胞の2次元(2D)、3次元(3D)イメージングにも成功したことから(図2)、動物細胞の場合には、PPK1遺伝子が極めて有効なレポーター遺伝子になり得ると考えられる(Biotechniques, vol.42, 209-215, 2007)。
図2. 31P-MRIを用いた動物細胞における遺伝子発現の非侵襲的・定量的な可視化(2D-, 3D-imaging).
|
| 分子細胞医科学研究室<古久保・高井グループ>では、転写・遺伝子発現等の基本的な生命現象に強い興味を持つ大学院生(博士前期課程、後期課程)を募集しています。当研究室の研究内容、教育方針、受験方法等についてもっと詳しく知りたいと思われた方は、気軽に下記宛メールで御連絡下さい。随時研究室見学、面談に応じています。
古久保 哲朗 <kokubo@tsurumi.yokohama-cu.ac.jp> |
|


 バフンウニ(Hemicentrotus
pulcherrimus)
バフンウニ(Hemicentrotus
pulcherrimus) バフンウニの卵を採取しているところ
バフンウニの卵を採取しているところ